|
|
BIOLOGÍA de 2º de BACHILLERATO |
||||||||||||||||||||||||||||||||||||||||||||||
|
|
tema II-4 - SISTEMAS DE MEMBRANAS |
||||||||||||||||||||||||||||||||||||||||||||||
|
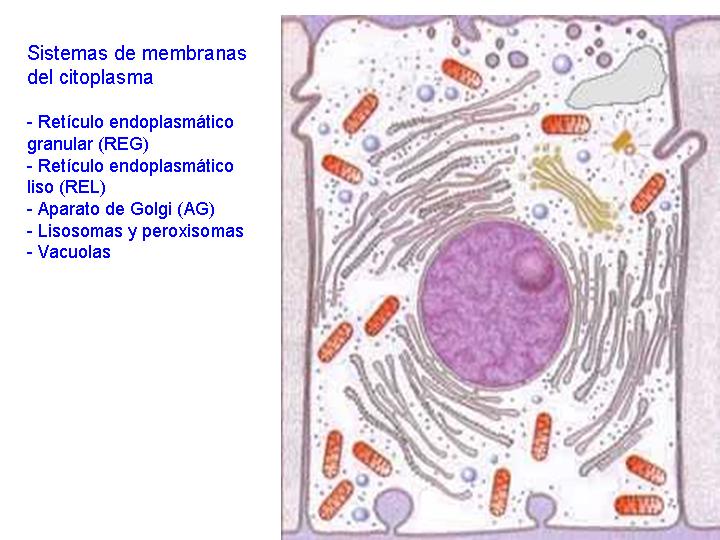
4) SISTEMAS DE MEMBRANAS DEL CITOPLASMA
El citoplasma se encuentra compartimentado por un complejo sistema de estructuras formadas por membranas biológicas relacionadas entre sí tanto físicamente como por la función que realizan, por lo que las estudiaremos conjuntamente.
Estos orgánulos son:
- Retículo endoplasmático granular (REG) - Retículo endoplasmático liso (REL) - Aparato de Golgi (AG) - Lisosomas y peroxisomas - Vacuolas
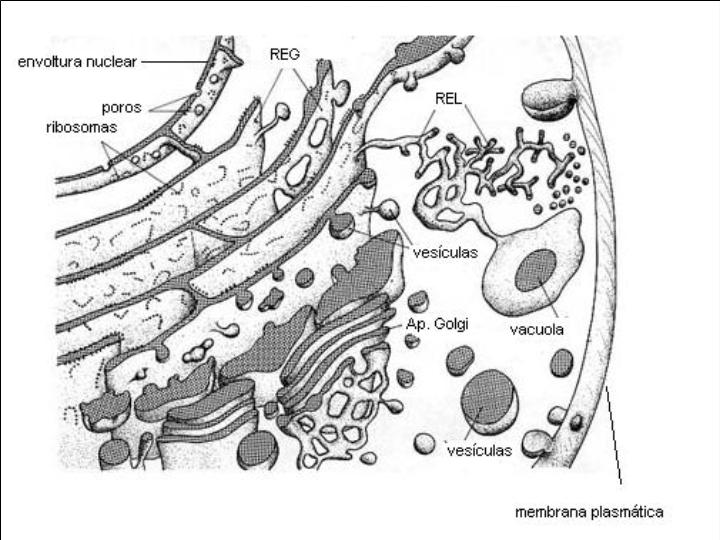
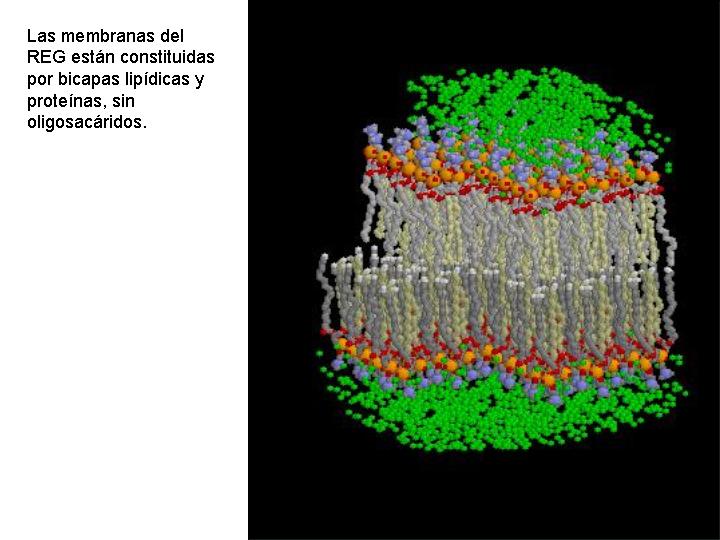
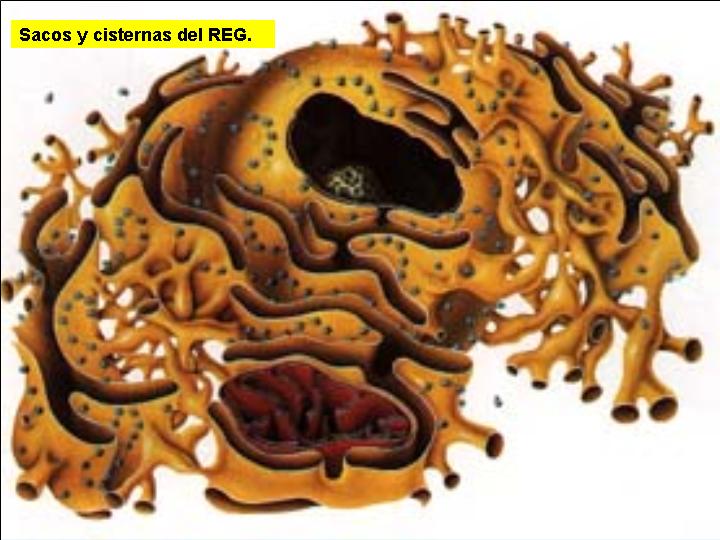
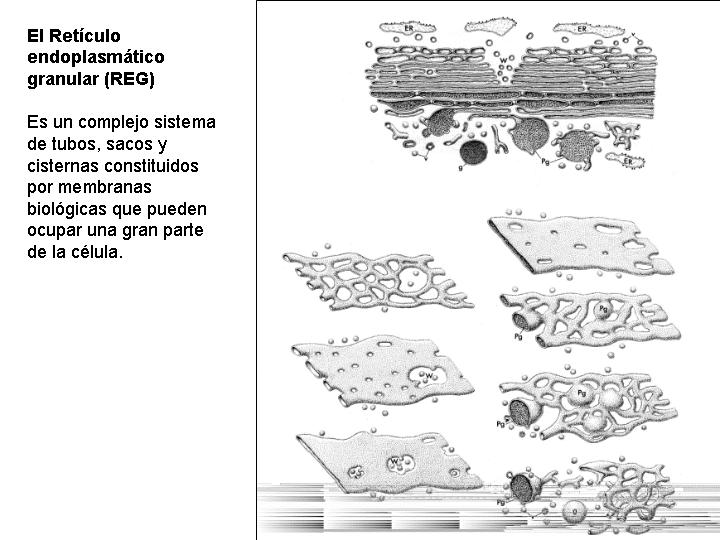
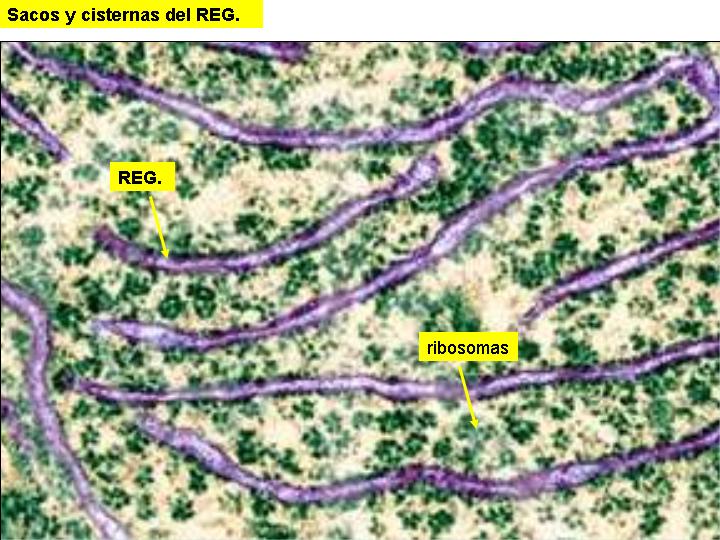
Es un complejo sistema de tubos, sacos y cisternas constituidos por membranas biológicas y que pueden ocupar una gran parte de la célula.
Las estructuras que forman el retículo se disponen, generalmente, en capas concéntricas paralelas al núcleo celular (como las hojas del bulbo de una cebolla). Es de destacar, que la envoltura nuclear es en realidad una estructura derivada del retículo endoplasmático.
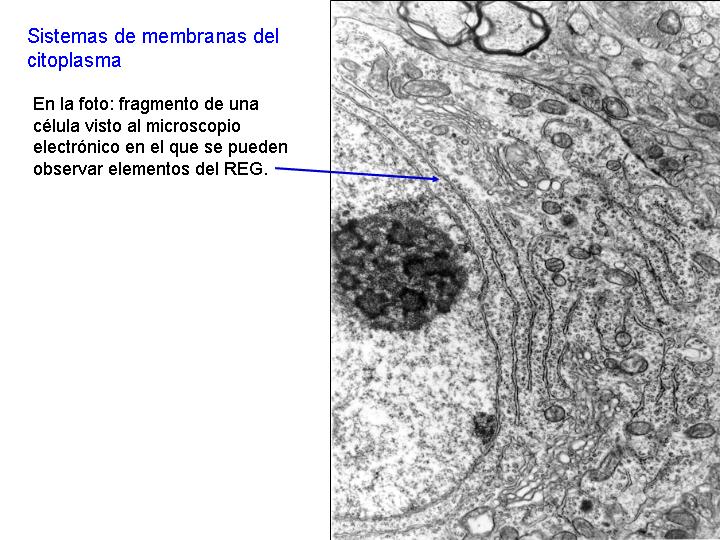
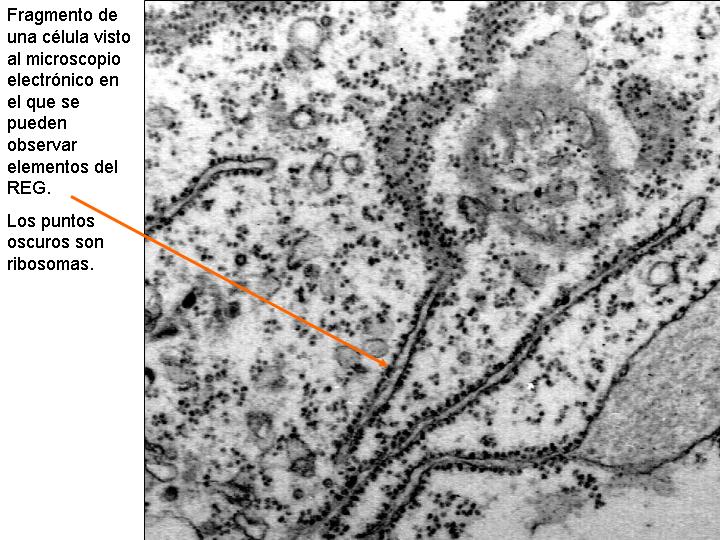
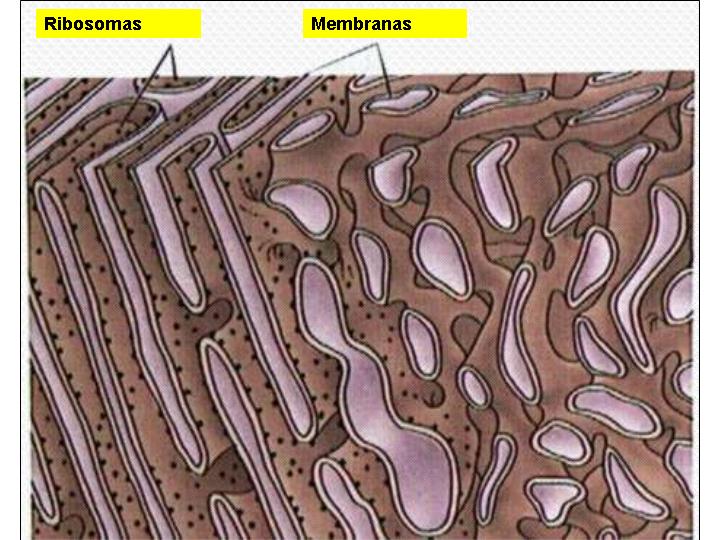
Existen dos tipos de retículo endoplasmático: el retículo endoplasmático liso (REL) y el retículo endoplasmático rugoso o granular (REG). En el REG se observan adheridos a las membranas unos gránulos: los ribosomas. En el REL no existen éstos gránulos y sus estructuras tienen formas más tubulares. También se diferencian en la función.
El retículo endoplasmático granular (REG) está muy desarrollado en las células que por su función deben de realizar una activa labor de síntesis, como es el caso de las células del páncreas y las células hepáticas. Si un animal es sometido a un ayuno prolongado, el REG de sus células pancreáticas se reduce considerablemente. Por el contrario, si se le suministra una rica dieta alimenticia, el REG se recupera. Esta recuperación se realiza a partir de zonas próximas a la envoltura nuclear.
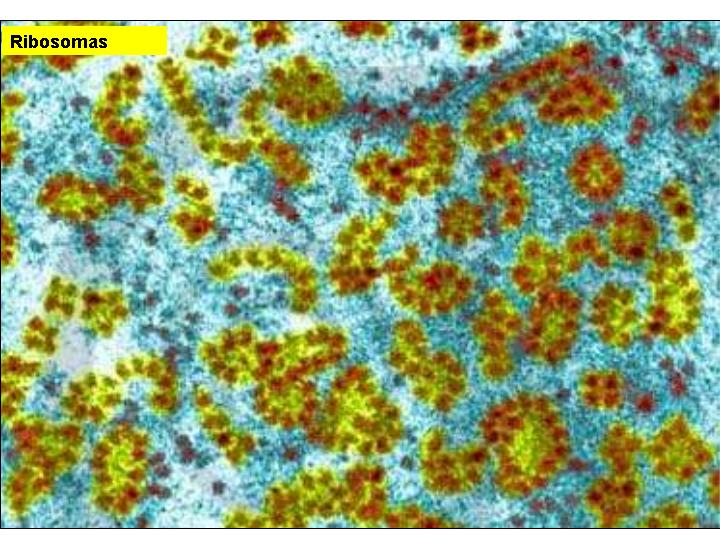
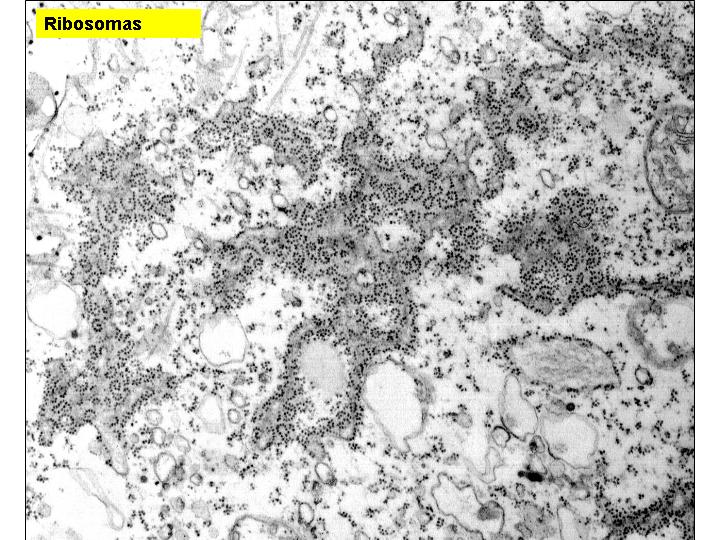
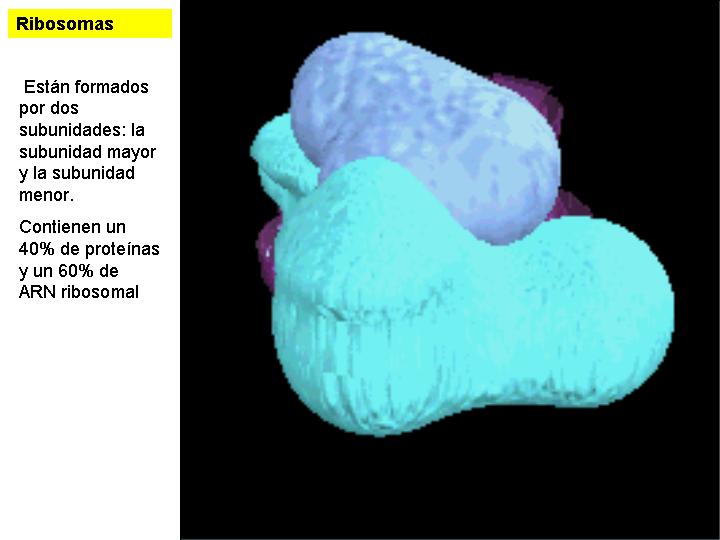
Son pequeños orgánulos invisibles al microscopio óptico y poco visibles al electrónico, no pudiéndose casi ni adivinar su estructura. Invaden en gran número el citoplasma y pueden estar libres o adheridos a las membranas del retículo endoplasmático granular. Los que están adheridos al REG intervienen en la síntesis de las proteínas de las membranas o de aquellas destinadas al exterior. Los ribosomas están constituidos básicamente por proteínas y ARN-r (40% de proteínas y 60% de ARN ribosomal).
Están formados por dos subunidades: la subunidad mayor y la subunidad menor. En el citoplasma ambas están separadas pero pueden volver a unirse en el momento de la síntesis de proteínas.
EL APARATO DE GOLGI (AG)
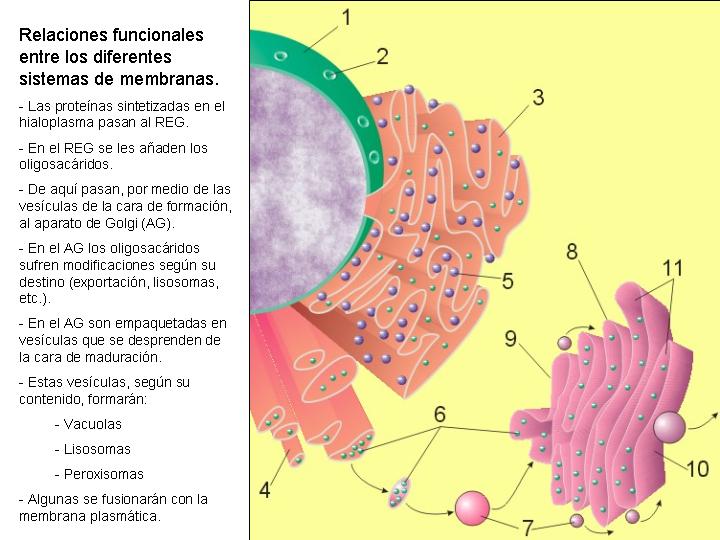
Está formado por unos conjuntos de sacos concéntricos muy apretados, mucho más concentrados y de menor tamaño que los del retículo endoplasmático granular y sin ribosomas. Cada conjunto de sacos es un dictiosoma. El número de dictiosomas por célula varía entre 5 ó 6 a algunas decenas, en función del tipo de célula y de su estado funcional. Todos ellos se encuentran relacionados física y funcionalmente.
Los dictiosomas presentan dos caras: una convexa, la cara de formación, y otra cóncava, la cara de maduración. De esta última se van desprendiendo pequeñas vacuolas que se independizan y que reciben el nombre de vesículas de secreción.
El AG se encuentra en permanente transformación. Sus sáculos se forman de manera continua por su cara de formación a partir de vesículas que se desprenden del REG y se desintegran por la cara de maduración para formar las vesículas de secreción.
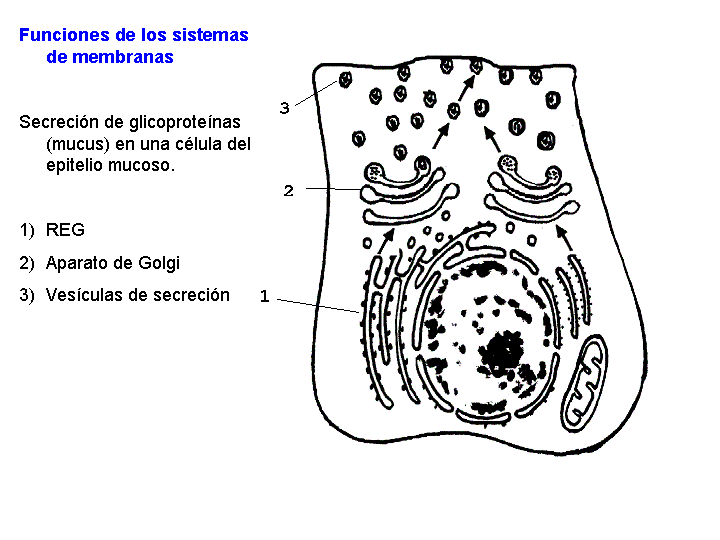
El aparato de Golgi se encuentra muy desarrollado en las células que realizan funciones de secreción, como las células secretoras de mucus del epitelio intestinal. Los dictiosomas son el sistema de transformación, embalaje y empaquetamiento de ciertas sustancias químicas, sobre todo de proteínas, para su almacenamiento o secreción.
LOS LISOSOMAS
Los lisosomas son pequeñas vesículas constituidas por membranas provenientes de los sistemas de membranas (AG y, ocasionalmente, REG). Se caracterizan por tener en su interior enzimas hidrolíticas, enzimas que rompen los enlaces de los polímeros por adición de H2O. Estas enzimas están empaquetadas e inactivas en los lisosomas y así se evita que puedan destruir las propias estructuras celulares.
Los lisosomas se originan en los dictiosomas del aparato de Golgi y, en algunos casos, en ciertas regiones del retículo endoplasmático granular a partir de vesículas que se destacan de los sáculos de los dictiosomas. Sólo están presentes en las células animales.
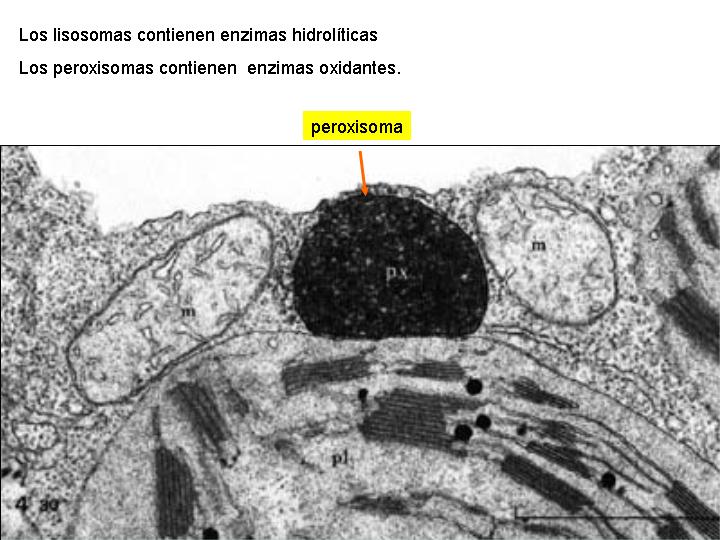
Son parecidos a los lisosomas, diferenciándose de estos en que contienen enzimas que degradan los ácidos grasos y los aminoácidos. Como estos procesos generan peróxidos, contienen también catalasa, enzima que descompone los peróxidos y en particular el H2O2 en H2O y O2.
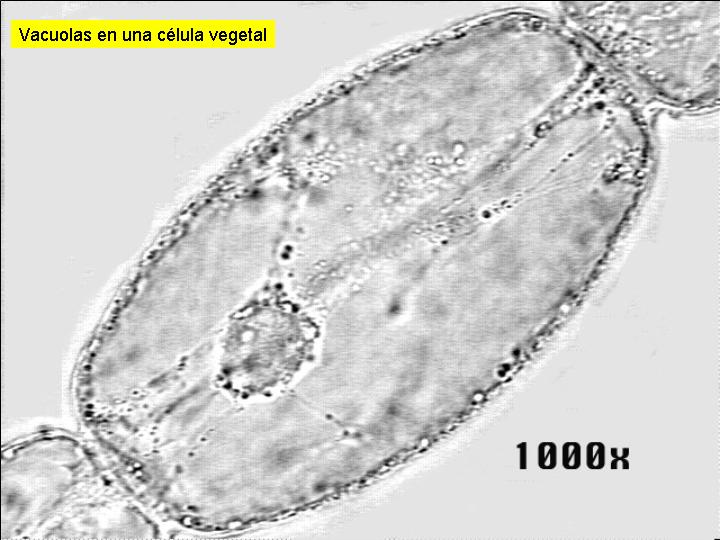
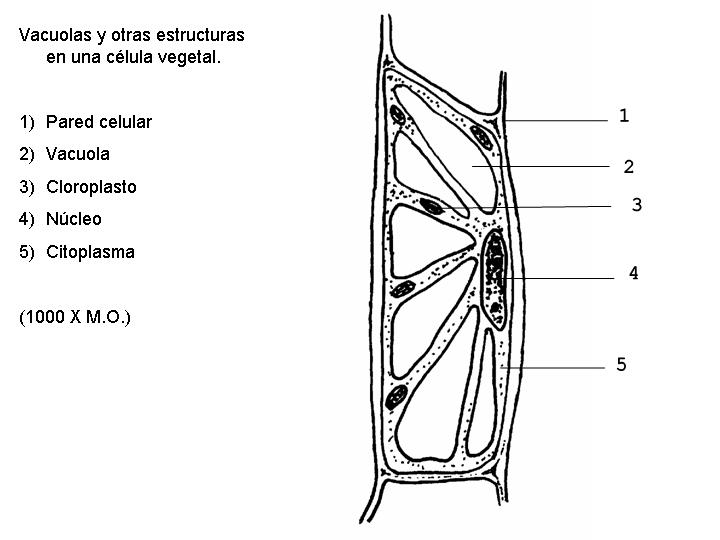
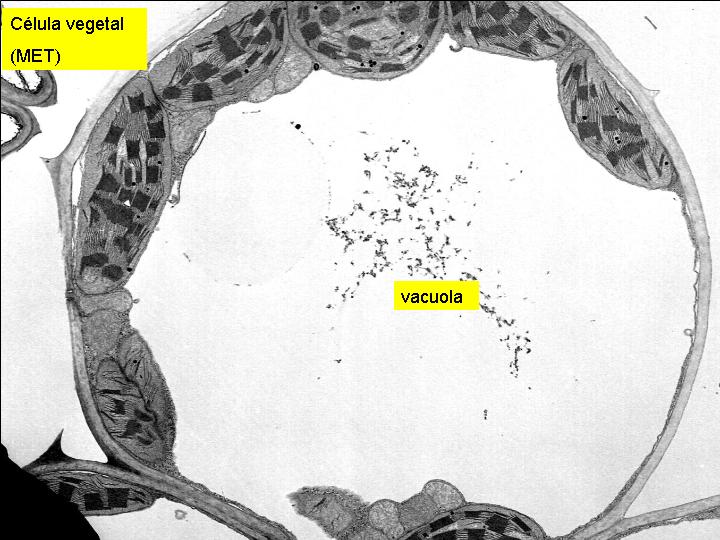
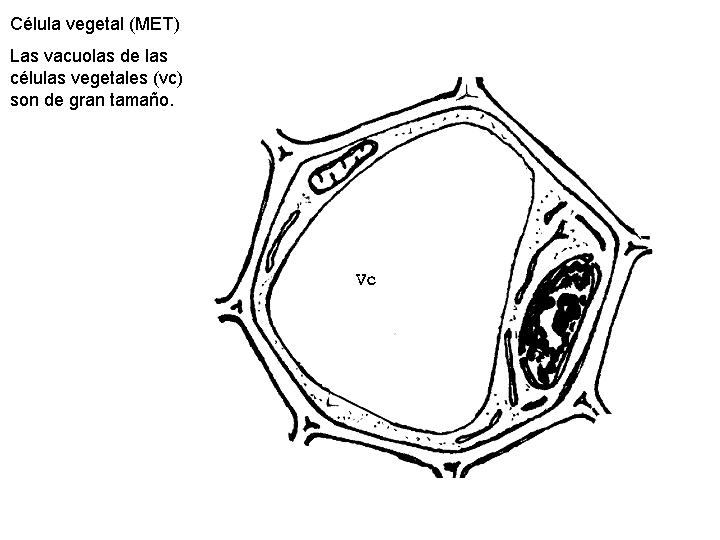
Son estructuras celulares variables en número y forma. En general están constituidas por una membrana y un contenido interno. Hay diferencias entre las vacuolas de las células vegetales y las de las células animales. Las células vegetales es frecuente que presenten una única o unas pocas vacuolas de gran tamaño. Las células animales, en el caso de tener vacuolas, son de pequeño tamaño.
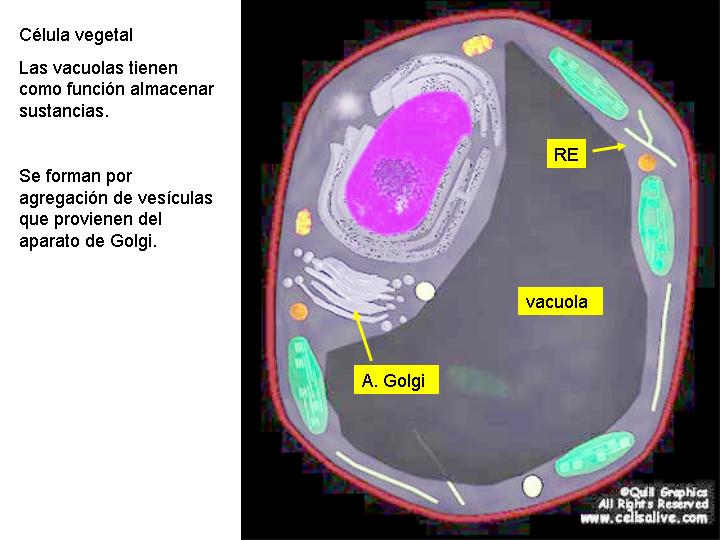
Las vacuolas se originan por la agregación de las pequeñas vesículas formadas a partir de los dictiosomas de aparato de Golgi o por invaginación de la membrana plasmática (endocitosis).
Las vacuolas, en general, tienen función de almacenamiento de sustancias de reserva y, en ciertos casos, de almacenamiento de sustancias tóxicas.
Existen otras estructuras que se llaman también vacuolas pero cuya función es muy diferente. Así:
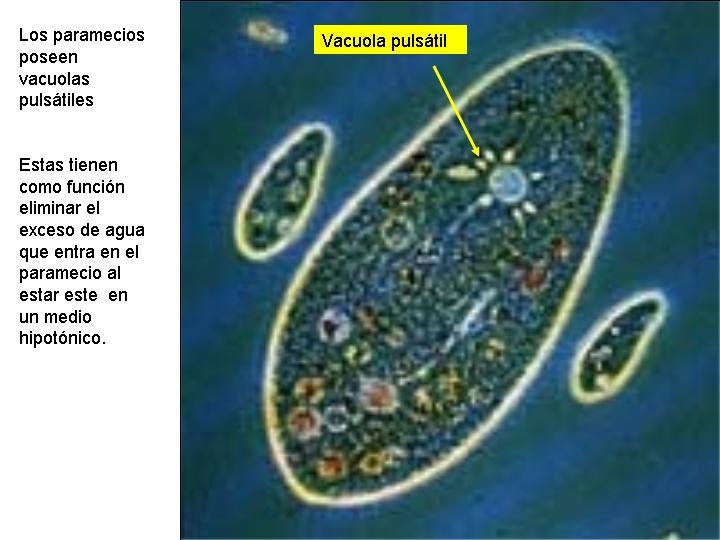
- Las vacuolas pulsátiles, como las que se observan en muchos organismos unicelulares de las aguas dulces, por ejemplo, el paramecio. Este organismo, al vivir en agua dulce, su citoplasma es hipertónico con respecto al exterior, por lo que se produce una entrada continua de agua. Las vacuolas pulsátiles extraen el agua del citoplasma y la expulsan al exterior por tansporte activo.
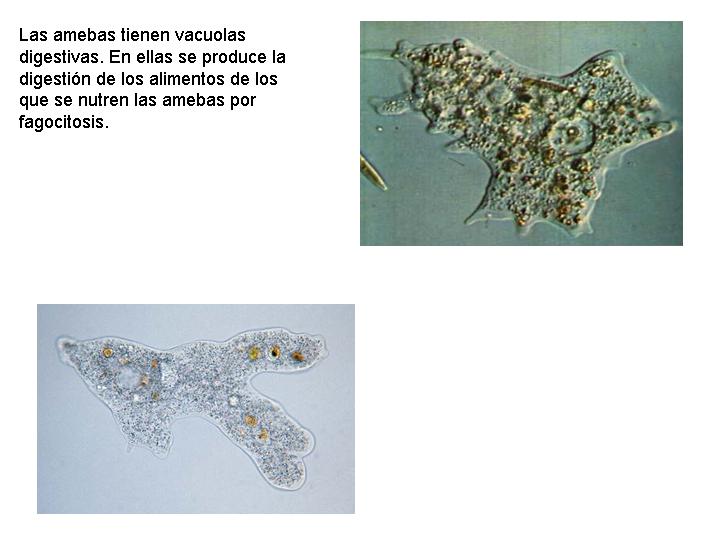
- Las vacuolas digestivas. Se dan en las células que capturan alimentos del medio y los engloban en una membrana formando una vacuola llamada vacuola digestiva. En esta vacuola es donde se va a producir la digestión de esas sustancias nutritivas. Una vez digeridas pasan al interior de la célula y los productos de desecho son eliminados hacia el exterior.
FUNCIONES DE LOS SISTEMAS DE MEMBRANAS
1) SÍNTESIS DE GLICOPROTEÍNAS
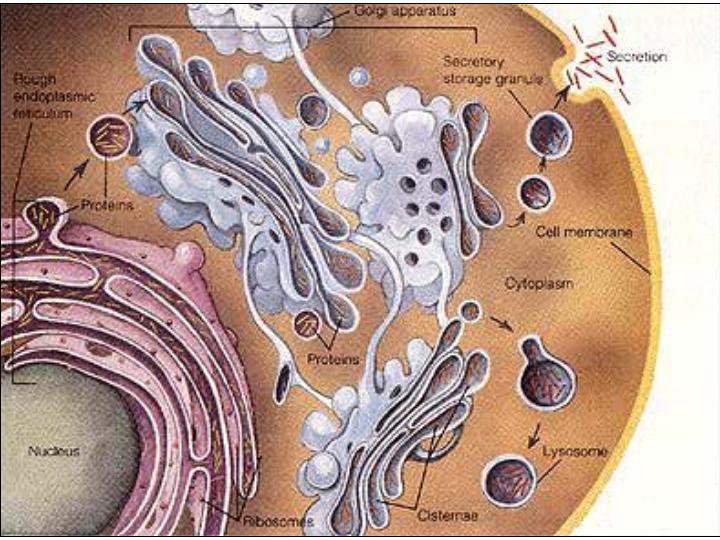
Los sistemas de membranas están relacionados con la síntesis, maduración y transporte de proteínas y glicoproteínas. Intervienen, sobre todo, en los procesos subsiguientes a la síntesis de las proteínas de secreción, las proteínas de las membranas y las enzimas de los lisosomas.
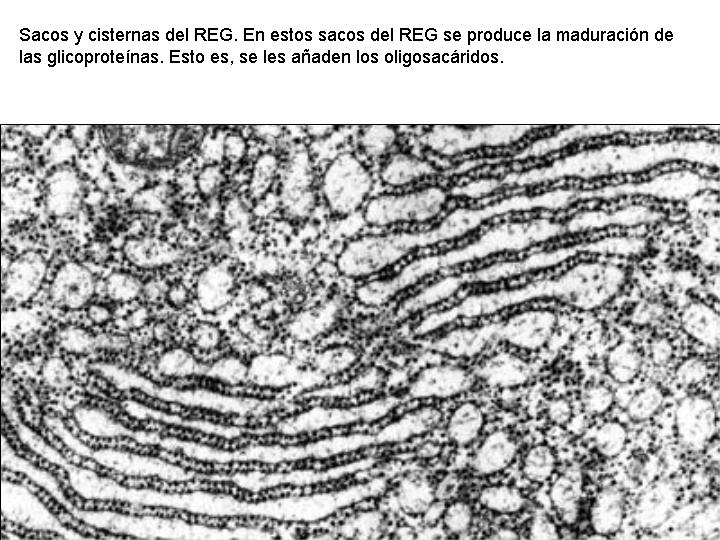
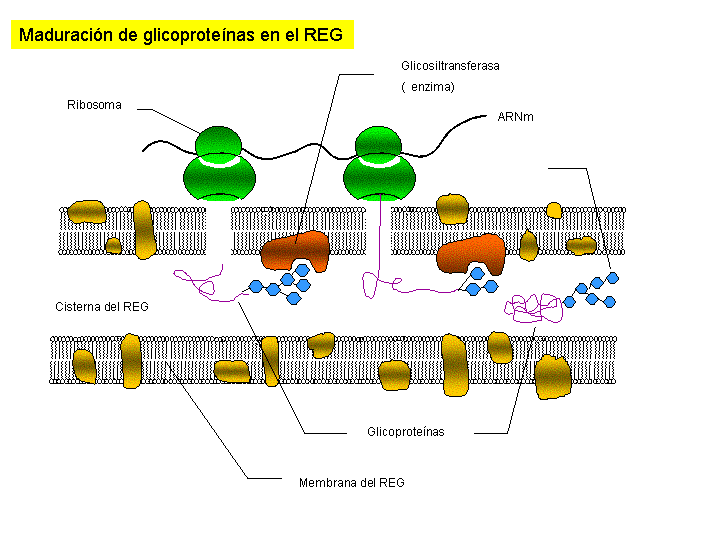
Las proteínas de secreción, proteínas destinadas a ser exportadas por la célula, y las proteínas de membrana se sintetizan en el citoplasma en regiones próximas al REG, esta es la razón por la que los ribosomas se encuentran fijos a las membranas del REG. Estas proteínas, según se van sintetizando, atraviesan la membrana de REG y pasan a su interior. Una vez en el interior del REG, en el caso de las glicoproteínas se va a producir la síntesis de los oligosacáridos. Los oligosacáridos de las membranas son relativamente sencillos y todos ellos son variaciones sobre dos únicos modelos. Uno de ellos es sintetizado en el REG y de aquí pasa al AG por medio de las vesículas que se unen a su cara de formación. En el AG los oligosacáridos sufren la adición o eliminación de determinados monosacáridos hasta su configuración definitiva. Existe otro tipo de oligosacáridos que es sintetizado totalmente en los sáculos del AG.
Las enzimas de los lisosomas son también glicoproteínas que se forman en el REG y en el AG. Una vez separadas las vesículas de los dictiosomas, su pH interno se vuelve ácido (pH=4,8) y las enzimas se sueltan de la membrana quedando libres en el interior del lisosoma. Las enzimas de los lisosomas en estas condiciones se encuentran inactivas, en tanto en cuanto el lisosoma no entre en funcionamiento.
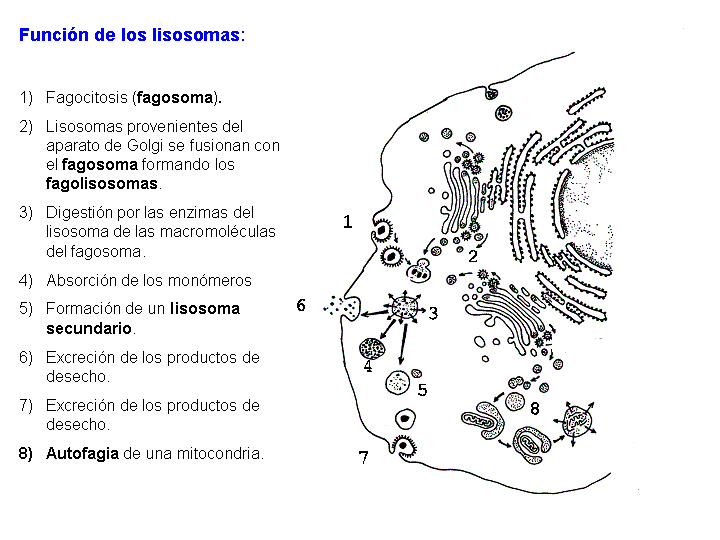
Función de los lisosomas: Hemos visto que ciertas células tienen la capacidad de ingerir sustancias por medio de fenómenos de endocitosis. Las sustancias son englobadas por la membrana plasmática que, a continuación, se invagina formando una vesícula denominada fagosoma. El fagosoma se fusiona con los lisosomas formando los fagolisosomas. Las grandes moléculas contenidas en el fagosoma: polisacáridos, proteínas, ácidos nucleicos, etc., son sometidas a la acción del medio ácido de los lisosomas y a las enzimas, que en este momento ya son activas. Los polímeros son hidrolizados y transformados en moléculas menores: monosacáridos, aminoácidos, etc., que se difunden a través de la membrana hacia el citoplasma. Quedan en el lisosoma los productos no degradados. Un lisosoma que ya ha actuado recibe el nombre de lisosoma secundario y conserva aún la capacidad de unirse a nuevos fagosomas. Las sustancias no degradadas se van acumulando progresivamente en el interior de los lisosomas secundarios. En ciertos organismos, estos lisosomas secundarios pueden fusionarse con la membrana plasmática y expulsar su contenido al exterior (exocitosis). En los organismos pluricelulares lo normal es que los lisosomas secundarios se transformen en cuerpos residuales. Esta acumulación de cuerpos residuales en una célula a lo largo de su vida es un signo de degeneración celular.
La membrana de los lisosomas puede englobar también orgánulos celulares que de esta manera son digeridos. Por este sistema la célula renueva sus estructuras celulares.
2) SÍNTESIS DE LOS POLISACÁRIDOS DE LA PARED CELULAR
En el aparato de Golgi se produce la polimerización de los polisacáridos y, en particular, la síntesis de la celulosa que constituye la sustancia fundamental de las paredes de las células vegetales.
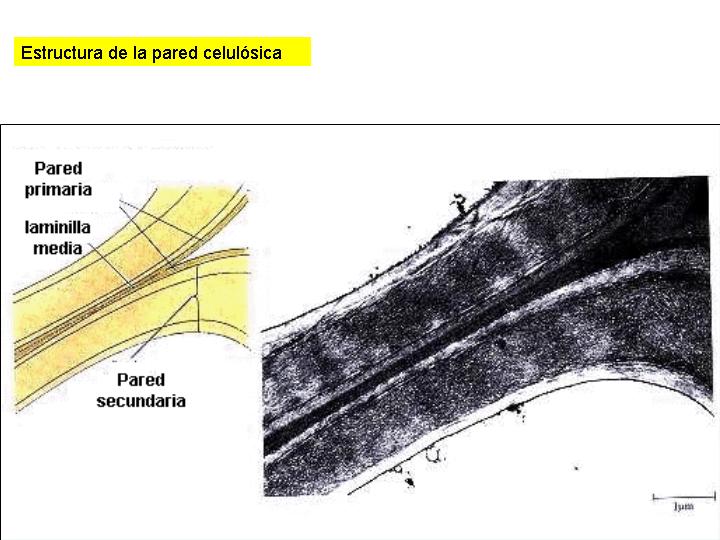
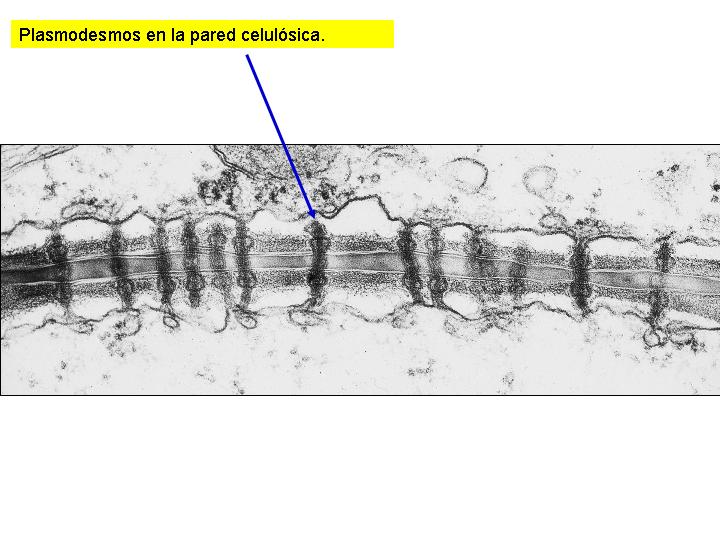
Las células vegetales disponen de una estructura que las envuelve denominada pared celular, constituida, fundamentalmente, por celulosa. La celulosa está formada por moléculas de glucosa unidas entre sí mediante enlaces ß (1-4). Esto hace que las moléculas de celulosa adopten una conformación lineal y que se puedan establecer puentes de hidrógeno entre moléculas dispuestas en paralelo formando microfibrillas entre las que se sitúan entrecruzadas moléculas de otras sustancias, como la lignina, que le da a la pared una gran rigidez, o ceras, que la impermeabilizan. La pared celular no es un orgánulo celular sino un producto de secreción de la célula que deposita en su exterior estas sustancias concéntricamente. La pared celular aparece además atravesada por una gran cantidad, hasta 20 000 en ciertos casos, de finísimos conductos denominados plasmodesmos. Los plasmodesmos comunican el protoplasma de las células contiguas que, en cierto modo, forman una unidad.
3) FUNCIONES DEL RETÍCULO ENDOPLASMÁTICO LISO (REL)
El retículo endoplasmático liso está relacionado con el metabolismo (síntesis, degradación y transporte) de los lípidos. Las hormonas esteroídicas son sintetizadas en el REL. Se ha observado que también interviene en los procesos para metabolizar ciertos medicamentos y determinadas sustancias tóxicas.
|
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||